
Uno de los objetivos principales de la Unidad de Entomología del IVIA es la búsqueda y desarrollo de nuevas estrategias de control sostenible de plagas y enfermedades en cultivos hortícolas. Recientemente, se ha demostrado cómo la exposición de plantas de pimiento a ciertos volátiles orgánicos induce respuestas defensivas que se traducen en una mayor resiliencia frente a herbívoros plaga. En este trabajo se resume cómo se han alcanzado estos resultados y se discute la posibilidad de utilización práctica de esta nueva herramienta de control basada en la comunicación entre plantas.
Chaymaa Riahi, Miquel Alonso-Valiente, Alberto Urbaneja y Meritxell Pérez-Hedo. Instituto Valenciano de Investigaciones Agrarias (IVIA). Centro de Protección Vegetal y Biotecnología, (IVIA), Valencia.
El pimiento es uno de los cultivos estratégicos en la cuenca mediterránea y en particular en el sureste español. A principios del presente siglo, la protección fitosanitaria de este cultivo en España evolucionó de un uso exclusivo e intensivo de insecticidas de origen químico a una gestión basada principalmente en estrategias de control biológico de plagas. Desde los primeros éxitos en invernaderos del Campo de Cartagena, las superficies de producción del pimiento empleando sistemas de control biológico han aumentado considerablemente, siendo en la actualidad este cultivo el ejemplo paradigmático del empleo de agentes de control biológico (van Lenteren et al., 2018).

Figura 1. Plagas más importante del cultivo de pimiento.
Las plagas que afectan a la producción del cultivo de pimiento tanto en invernadero como al aire libre son numerosas (figura 1). El trips occidental de las flores Frankliniella occidentalis (Pergande) (Thysanoptera: Tripidae) se considera la plaga clave de este cultivo, no solamente por el daño directo que produce alimentándose de la planta, sino también por su capacidad de transmisión del virus del bronceado del tomate (TSWV). Además, varias especies de áfidos perjudican la producción del pimiento. Principalmente, Myzus persicae (Sulzer), Myzus persicae nicotianae Blackman, Aphis gossypii Glover, Macrosiphum euphorbiae Thomas y Aulacorthum solani Kaltenbach (Hemiptera: Aphididae) ocasionan graves daños que se caracterizan por el amarillamiento y el debilitamiento de las plantas y el favorecimiento de la colonización de hongos por la gran cantidad de melaza secretada. Igualmente, el pulgón M. persicae actúa como vector del virus Y de la patata (PVY) y el virus del pepino (CMV).
Aunque el pimiento no es el huésped preferido de las moscas blancas (Trialeurodes vaporariorum Westwood y Bemisia tabaci Gennadius –Hemiptera: Aleyrodidae–), cuando las condiciones lo permiten, esta plaga es capaz de desarrollarse a la perfección. Debido a la extracción de la savia y la secreción de la melaza, la mosca blanca provoca la aparición de negrilla en hojas y fruto, lo cual debilita la planta causando un marchitamiento general y en consecuencia, reduce la calidad del fruto. Otras plagas segundarias del pimiento, que en ocasiones pasan a ser importantes, son la araña roja Tetranychus urticae Koch (Acari: Tetranychidae) y la rosquilla verde Spodoptera exigua Hübner (Lepidoptera: Noctuidae) que provocan necrosis y defoliación, respectivamente, impidiendo el óptimo desarrollo de la planta.
Control biológico en pimiento
El establecimiento de programas de gestión de plagas basados en control biológico ha supuesto una clara mejoría en el control de plagas en cultivos hortícolas de la cuenca mediterránea (van Lenteren et al., 2020). Estos programas han limitado al máximo el uso de plaguicidas, lo que ha satisfecho enormemente las exigencias del mercado en lo que a la no presencia de residuos de plaguicidas se refiere (van Lenteren et al., 2018). El primer paso fue frenar el impacto de las dos plagas clave en la mayoría de los cultivos hortícolas, el trips (F. occidentalis) y las moscas blancas, plagas que habían adquirido resistencias a muchos de los productos fitosanitarios aplicados en el siglo pasado.
En pimiento, el uso combinado de dos depredadores polífagos ha resultado ser la clave del éxito de la gestión de estas dos importantes plagas: el chinche depredador Orius laevigatus (Fieber) (Hemiptera: Anthocoridae) y el ácaro depredador Ambyliseius swirskii Athias-Henriot (Acari: Phytoseiidae) (Calvo et al., 2015; Sánchez y Lacasa, 2002). El uso de estos dos depredadores polífagos junto con el empleo de otros enemigos naturales y el uso de plaguicidas selectivos ha permitido mantener a raya la mayor parte de fitófagos plaga en el cultivo del pimiento.

Foto 1. Exposición de las plantas de pimiento a volátiles de plantas (HIPVs) mediante la utilización de un difusor polimérico que permite una liberación del volátil constante y duradera (Pérez-Hedo et al., 2021d).
Sin embargo, los pulgones escapan a veces a este control, siendo necesario o bien el empleo de gran cantidad de enemigos naturales (tanto parasitoides como depredadores) que encarecen el precio del programa de control o bien el uso de plaguicidas que pueden romper el equilibrio biológico (Belliure et al., 2008). Por ello, se han buscado alternativas para dotar al cultivo de pimiento del sureste español de nuevas estrategias de control biológico para el control de pulgones (Pérez-Hedo y Urbaneja, 2015; Pineda y Marcos-García, 2008).
Una de las opciones que se ha barajado como alternativa factible para el control de pulgones es el uso de míridos depredadores zoofitófagos que se alimentan tanto de plantas como de insectos herbívoros (Pérez‐Hedo et al., 2021a). Además, se ha comprobado cómo los míridos Nesidiocoris tenuis (Reuter) y Macrolophus pygmaeus Rambur (Hemiptera: Miridae) son compatibles con el uso de A. swirskii y pueden llegar no solo a controlar los pulgones sino también a regular las poblaciones de trips y moscas blancas (Bouagga et al., 2018a; Pérez-Hedo y Urbaneja, 2015).
Además de la depredación directa sobre insectos herbívoros, los míridos depredadores presentan un valor añadido para su utilización en pimiento y es su capacidad de activar las defensas de la planta, a través de su fitofagia. Trabajos de nuestro grupo han demostrado que tanto N. tenuis como M. pygmaeus, así como O. laevigatus, cuando se alimentan de la planta de pimiento e incluso al ovipositar, inducen respuestas defensivas en la planta (Bouagga et al., 2020, 2018c, 2018b). Esta inducción de defensas se ha comprobado cómo infiere a las plantas una mayor resistencia a varias especies plaga y les hacen más atrayentes frente a enemigos naturales.
Comunicación entre plantas
Cuando las plantas son atacadas por herbívoros, estas activan mecanismos de defensa que implican diferentes vías de transducción reguladas por fitohormonas, como el ácido salicílico (SA), ácido jasmónico (JA), ácido abscísico (ABA) y etileno (ET). La fitofagia estimula la liberación de una mezcla de los volátiles de planta HIPVs dominada por ésteres, terpenos, aldehídos, alcoholes y compuestos aromáticos, y que indirectamente influye en las plagas atrayendo a sus depredadores y/o parasitoides. Los compuestos volátiles también participan en muchas interacciones multitróficas incluyendo la señalización intra-planta, la interacción flor-polinizador y los enemigos naturales de los herbívoros (Arimura et al., 2009).
El sistema de comunicación de las plantas está basado en el uso de compuestos orgánicos volátiles. Esas mezclas químicas son interceptadas como señal de advertencia por plantas sanas que se encuentran cerca de las atacadas por fitófagos. Las plantas sanas que reciben estas señales son capaces de activar sus defensas para prevenir el inminente ataque que les acecha (Mauch-Mani et al., 2017). Por tanto, si pudiéramos conocer qué volátiles son los que activan defensas en plantas sanas, se podría explorar su uso como nueva herramienta de gestión de plagas (Pérez-Hedo et al., 2020).
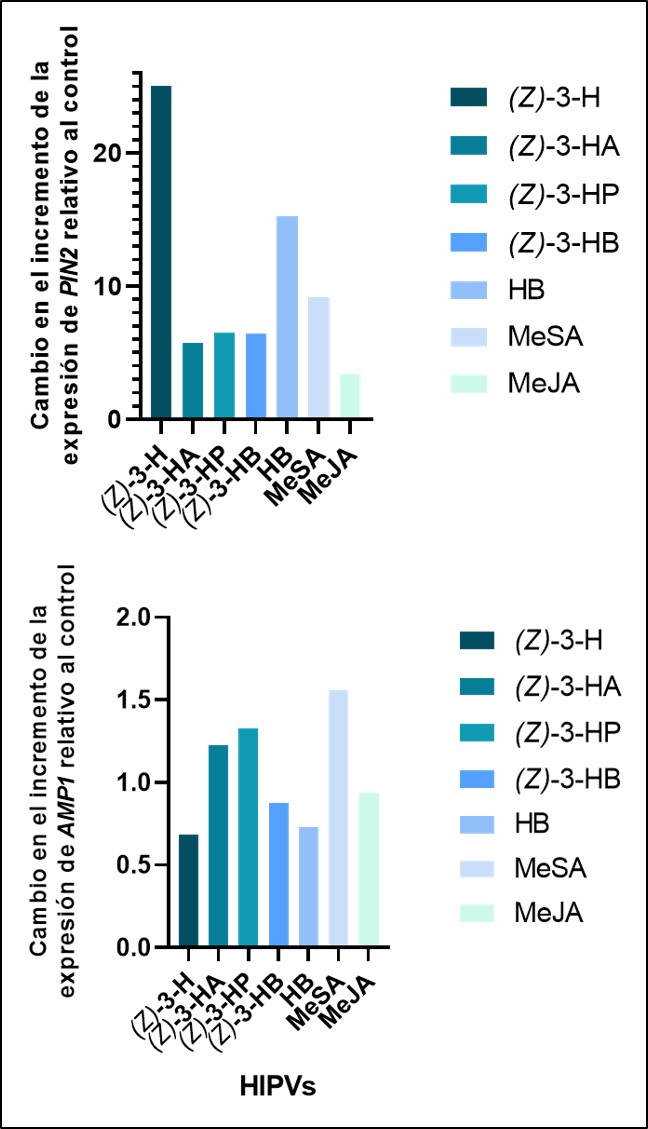
Figura 2. Cambio en el incremento (Fold Change) de los genes PIN2 y AMP1 marcadores de las vías de señalización del JA y SA cuando son expuestos a volátiles de planta respecto a un control sin exposición. Adaptado de Riahi et al. (2022).
Siguiendo esta línea, nuestro grupo demostró que, en plantas de tomate, la exposición por separado a siete volátiles de planta (1-hexanol, (Z)-3-hexenol, (Z)-3-hexenil acetato, (Z)-3-hexenil propanoato, (Z)-3-hexenil butanoato, hexil butanoato, metilo salicilato y metilo jasmonato) durante 24 horas fue capaz de activar la respuesta defensiva en las plantas de tomate y de sobreexpresar genes de defensa relacionados con las rutas metabólicas del JA, SA y el inhibidor de la proteína I de la planta (Meritxell Pérez-Hedo et al., 2021b). Además, se reveló el efecto de repelencia de dos volátiles potencialmente seleccionados [(Z)-3-hexenil propanoato y metilo salicilato] sobre las plagas del tomate F. occidentalis, B. tabaci y Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) y el efecto de atracción hacia el parasitoide de la mosca blanca Encarsia formosa (Gahan) (Hymenoptera: Aphelinidae) (Pérez-Hedo et al., 2021a, 2021b).
Exposición a volátiles en pimiento
Tras los resultados obtenidos en tomate, y conociendo que las plantas de pimiento también emitían un perfil de volátiles similar al de tomate cuando eran picadas por míridos zoofitófagos (Bouagga et al., 2018b), se decidió estudiar si las plantas de pimiento respondían de manera similar a la exposición de volátiles de planta. Para ello, se ensayaron los siguientes volátiles: (Z)-3-hexenol [(Z)-3-H], (Z)-3-hexenil acetato [(Z)-3-HA], (Z)-3-hexenil propanoato [(Z)-3-HP], (Z)-3-hexenil butanoato [(Z)-3-HB], hexil butanoato [HB], metilo salicilato [MeSA] y metilo jasmonate [MeJA] que fueron expuestos individualmente a plantas de pimiento mediante un emisor polimérico que permite una liberación constante y prolongada del volátil a utilizar (foto 1) (Riahi et al., 2022).
Mediante el análisis de la expresión genética se demostró que la expresión de los genes de defensa PIN2 y AMP1 marcadores de las rutas metabólicas de JA y SA respectivamente, fueron sobreexpresados (figura 2). Además, se estudió el comportamiento de respuesta de F. occidentalis en el análisis del estímulo olfativo en un olfactómetro en forma de tubo donde se observó que fue repelida por aquellas plantas expuestas al (Z)-3-hexenil propanoato y al metilo salicilato. Por otra parte, el chinche depredador O. laevigatus mostró una atracción a plantas de pimiento expuestas a (Z)-3-hexenol, (Z)-3-hexenil propanoato, (Z)-3-hexenil butanoato, metilo salicilato y metilo jasmonate. Estos resultados demuestran que los volátiles actúan como elicitores de las defensas de la planta de pimiento al mejorar las vías de señalización defensiva. Anticipamos que estos resultados son un punto de partida para la integración de enfoques basados en volátiles en los sistemas de gestión de plagas de pimiento, lo que pueda proporcionar una estrategia sostenible para mejorar la gestión de plagas.
Conclusión
El uso de los volátiles de planta podría resultar en un nuevo enfoque para un manejo respetuoso para el medio ambiente y con garantía sanitaria para el control de plagas al aprovechar el potencial de la comunicación entre las plantas y aprovechar así sus mecanismos de defensas. En este contexto, en la actualidad se está ensayando el efecto sobre el desarrollo y reproducción de varias plagas sobre plantas expuestas a la emisión de volátiles.












